ОТДЕЛ MYXOMYCOTA = EUMYCETOZOA
Миксомице́ты (лат. Mycetozoa, от греч. μύκητος — «гриб» и ζωον — «животное» или Myxomycota от μύξα — «слизь» и μύκητος) — тип слизевиков (грибоподобных организмов), входящий в состав супергруппы Amoebozoa. Насчитывает около 1000 видов. На определённой стадии жизненного цикла все миксомицеты имеют вид плазмодия или псевдоплазмодия.
 |
Классы Myxomycetes и Protosteliomycetes, имеющие плазмодий, относят к плазмодиальным (неклеточным) слизевикам, а Dictyosteliomycetes, которые имеют псевдоплазмодий — к клеточным. У большинства видов он виден невооружённым глазом и способен двигаться. Из плазмодия или псевдоплазмодия формируются спороношения, которые часто напоминают внешним видом плодовые тела грибов.
Споры прорастают подвижными клетками — зооспорами или миксамёбами, из которых различными путями образуется плазмодий или псевдоплазмодий.
|
Слизевики — гетеротрофы. Зооспоры, миксамёбы и плазмодии способны питаться осмотрофно (всасыванием питательных веществ через клеточную мембрану) и (или) путём эндоцитоза (захватывать внутрь клетки пузырьки с частицами пищи).
Все миксомицеты свободноживущие и живут в наземных местообитаниях.
Классификация
Ведущим методом в систематике миксомицетов остаётся морфологический анализ признаков спороносных структур. Также вопросы их филогении и систематики решают молекулярно-биологическими и электронно-микроскопическими исследованиями (Spiegel et al., 1995; Lutzoni et al., 2004; Fiore-Donno et al., 2005).
Показано, что признаки ультраструктуры жгутикового аппарата зооспор Myxomycetes дают возможность для выяснения их родственных отношений с церкомонадами и протостелидами. Исследования ультраструктуры зооспор миксомицетов из различных порядков указывают на значительную консервативность строения правого и дорсального корешков жгутика и значительную изменчивость в строении левого корешка и фибриллярных связок в основании жгутика (Karpov et al., 2003). Сходство в строении корешковой системы жгутиков у гиперамёб, церкомонад и миксомицетов позволило некоторым авторам рассматривать их в качестве членов одной филогенетической ветви (Karpov, 1997). Однако эти данные противоречат гипотезе, основанной на молекулярных исследованиях, согласно которой миксомицеты близки к гиперамёбам, тогда как церкомонады отстоят от этой ветви довольно далеко (Zaman et al., 1999). Анализ нуклеотидных последовательностей малых субъединиц рибосомальной РНК показывают связь между миксомицетами и Hyperamoeba. Нуклеотидная последовательность малой субъединицы рибосомальной РНК у Hyperamoeba почти идентична таковой у миксомицета Stemonitis pallida и некоторых видов порядка Physarales (Walker et al., 2003).
В настоящее время к миксомицетам относят 3 класса: Dictyosteliomycetes (Диктиостелиомицеты); Myxomycetes (Собственно слизевики); Protosteliomycetes (Протостелиевые).
Протостелиевых с собственно миксомицетами сближают особенности ультраструктуры жгутикового аппарата зооспор, трубчатые кристы митохондрий, характер образования плодовых тел и строение плазмодиев. Видимо, это эволюционные ветви, отходящие от общего протостелиоподобного предка. Для диктиостелиевых (одной из групп «клеточных» слизевиков) показано, что наличие периферических ядрышек, находящихся в тесном контакте с ядерной мембраной, отсутствие жгутиковых стадий и крайне примитивный геном резко отделяют их от других слизевиков. Первые исследования генов большой и малой субъединиц рРНК (De Rijk et al., 1995) также указывали на то, что диктиостелиевые стоят далеко как от Myxomycetes, так и от лобозных амёб, из этих работ также следовал вывод о полифилии миксомицетов. |
|
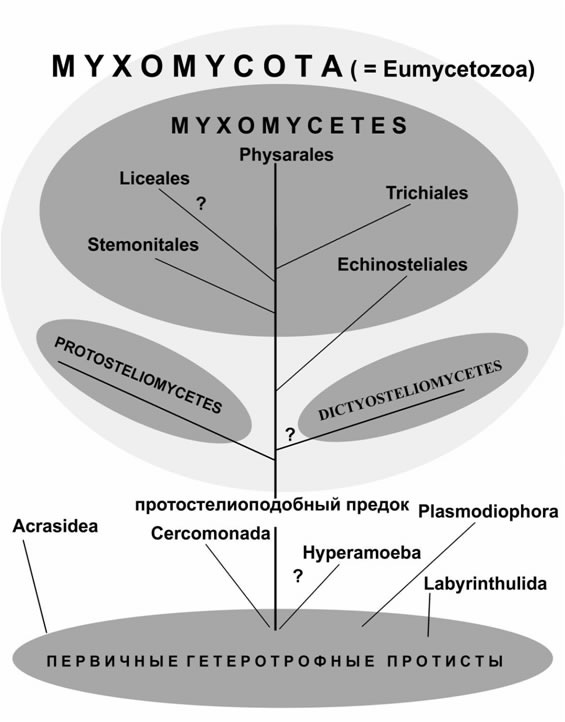
Однако недавние исследования, проведённые на основе анализа актина, α- и β-тубулина и фактора EF-1α, указывают на монофилию Dictyosteliomycetes, Protosteliomycetes и Myxomycetes и служат основанием рассматривать Mycetozoa как эволюционно целостную группу (Baldauf, 1999).
Строение
Вегетативное тело миксомицетов представлено плазмодием, то есть голой многоядерной клеткой. Это масса цитоплазмы с большим количеством (до нескольких миллионов) ядер, прозрачная или непрозрачная, бесцветная или жёлтая, красная, фиолетовая, чёрная и др.
У большинства видов плазмодий имеет вид сети из переплетающихся и сливающихся трубочек с веерообразным краем со стороны, куда он движется. Размеры плазмодия у разных видов различны. У некоторых он микроскопический, у некоторых — неограниченного размера и вырастает, как правило, до сантиметров или десятков сантиметров, иногда до метров. Плазмодий способен к активному амёбообразному движению: в направлении движения появляются протоплазменные выросты, а с противоположной стороны они втягиваются. Скорость движения плазмодия от 0,1 до 0,4 мм в минуту.
 |
Миксомицеты сапротрофны. Их плазмодии живут в тёмных сырых местах, богатых органическими веществами: в пустотах и расщелинах гнилых пней и колод, под корнями, опавшими перегнивающими листьями. В это время для плазмодия характерно движение к темноте, сырости и источникам пищи. Питается он, поглощая из субстрата питательные вещества, захватывая бактерии, микроскопических животных, споры грибов, и быстро увеличивается в размерах. После периода вегетативного роста, когда исчерпываются питательные вещества в окружающей среде, плазмодий выползает на свет, на поверхность пня или листовой подстилки. Здесь он образует спороношения, которые содержат огромное количество спор. Часто эти спороношения напоминают миниатюрные грибы, на основании чего миксомицеты традиционно относили к царству грибов (Kirk et al., 2001). Однако это сходство чисто поверхностное: миксомицеты отличаются от грибов рядом биохимических и ультраструктурных признаков, отсутствием клеточной стенки у вегетативного тела, характером питания и некоторыми другими особенностями. |
Процесс превращения плазмодия в спороношение в простейшем случае заключается в том, что плазмодий, не меняя формы, одевается перепончатой или хрящевой оболочкой, и внутри образуются споры.
Такое спороношение называется плазмодиокарп. Он похож на плоскую лепёшку или подушечку, иногда имеет неправильную форму. У многих видов спороношение более сложное: плазмодий разделяется на много частей и каждая превращается в спорангий. В состав спорангия входит округлое или вытянутое образование (споротека), содержащее споры и капиллиций — систему нитей в виде сети или каркаса. Споротека, как правило, покрыта оболочкой (перидием) и сидит на ножке. У некоторых видов спорангии на ранней стадии развития сливаются в единое образование, обычно довольно крупных размеров (до десятков сантиметров), одетое общей оболочкой. Такое спороношение называется эталием, а при неполном слиянии спорангиев — псевдоэталием. Для некоторых родов миксомицетов характерно присутствие в основании спороношений кожистых плёночек — гипоталлуса. При созревании спор оболочка спороношения (перидий) разрывается, и споры рассеиваются по воздуху. У многих миксомицетов распространению спор содействует капиллиций (система особых нитей, содержащаяся в спороношении); у разных миксомицетов он имеет различное строение и его признаки важны для систематики. |
Миксомицеты. Иллюстрация из книги «Kunstformen der Natur» Эрнста Геккеля (1904). Вверху — споры, зооспоры, плазмодий и миксамёбы, снизу — спороношения. |

Благодаря различным утолщениям на своей поверхности нити капиллиция способны к гигроскопическим движениям, при которых споровая масса разрыхляется и разбрасывается (Горленко и др., 1980).
Споры представляют собой микроскопически малые шаровидные клетки, бесцветные или окрашенные, одетые твёрдой оболочкой — гладкой или различным образом скульптурированной. Размеры, окраска и строение оболочки спор — это существенные признаки при определении миксомицетов, как и строение спороношения в целом.
В сухом виде споры могут сохраняться без изменений иногда в течение десятков лет, но, попав во влажный субстрат, быстро прорастают. При этом из разрыва их оболочки или, у некоторых видов, через поры выходят одна или несколько зооспор с двумя гладкими жгутиками неравной длины или амёбы (миксамёбы). Они могут превращаться друг в друга в зависимости от влажности и размножаться делением. Затем происходит половой процесс, то есть попарное слияние зооспор или миксамёб. В результате образуется зигота, из которой развивается новый плазмодий. Он опять уходит в тёмные и влажные места, и процесс начинается сначала.
Распространение
Многие виды миксомицетов являются космополитами. Однако некоторые из них, особенно виды, обитающие в тропиках и пустынях, имеют ограниченные ареалы. Наибольшее видовое богатство наблюдается в широколиственных лесах умеренной зоны и в мезоксерофильных лесах Средиземноморья. В умеренных поясах обоих полушарий миксомицеты чаще всего живут в лесах, встречаясь в виде плазмодиев или спороношений в течение лета, а иногда с ранней весны и до поздней осени. Но многие виды обнаруживают сезонность в своём развитии. Так, некоторые виды образуют спороношения только весной и прекращают спорообразование в середине лета, у других спорообразование начинается летом и продолжается вплоть до листопада. Многие виды миксомицетов обитают на крупных остатках древесины и опаде. Их плодовые тела обычно хорошо заметны в природе. Это во многом объясняет тот факт, что в большинстве микологических работ приводятся сведения главным образом о видах, связанных с этими субстратами. В то же время виды, обитающие на коре живых деревьев и на помёте растительноядных животных, часто имеют мелкие и плохо заметные в природе спорофоры и, как правило, не учитываются.
Значение
В последние десятилетия интерес к этим организмам возрос благодаря многочисленным цитологическим, биохимическим, биофизическим и генетическим исследованиям, в которых слизевики использовались как модельные организмы.
Изучение миксомицетов имеет большое значение для эволюционных и филогенетических построений, а также для систематики грибов и протистов (Кусакин, Дроздов, 1998; Карпов, 2000; Новожилов, Гудков, 2000).
Миксомицеты активно поедают бактерии, играя значительную роль в регуляции численности и состава бактериальной флоры почв (Madelin, 1983).
Таксономия миксомицетов базируется главным образом на морфологических признаках плодовых тел (спорофоров), которые относительно легко гербаризируются и поддаются количественному учёту. Присутствие вида в природе оценивается по наличию спорофоров на изучаемой площади или в образцах субстратов. Эти особенности миксомицетов и уникальная для протистов комбинация в жизненном цикле микроскопических амёб, плазмодиев и часто хорошо заметных спорофоров, позволяют использовать их в качестве модельного объекта для синэкологических и географических исследований.
Наряду с традиционным отбором спорофоров миксомицетов в полевых условиях, их можно получать в лаборатории, используя метод влажных камер. Его преимуществом является возможность легко стандартизировать условия культивирования и выявлять даже плохо заметные в природе виды в любое время года.
Систематика :